The model assumes that two independent genetic loci, a and b, are interacting in the following way (epistasis):
| locus a | locus b | phenotype | fitness |
| a0a0 | b0b0 | A0B0 | high |
| a0a0 | b1b1 | A0B1 | lower |
| a1a1 | b0b0 | A1B0 | lower |
| a1a1 | b1b1 | A1B1 | very high |
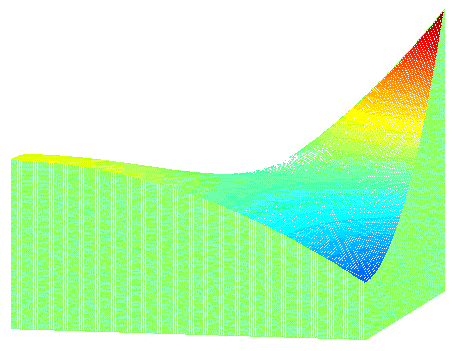
This can be illustrated as a fitness landscape, as shown below:
|
| Mean fitness of population (z-axis) as a function of the frequencies of gene a1 and b1 (x- and y-axes). |
The question is whether the population can move from the local maximum, represented by the low peak, through the valley to the global maximum, represented by the high peak in the fitness landscape.
The intuitive metaphor of a fitness landscape, as illustrated above, has misled previous theorists to believe that the trajectory from the low peak to the high peak goes through the saddle point (e.g. Barton & Rouhani 1987). My simulations show, however, that such a trajectory is extremely unlikely for lack of genetic variation. In stead the trajectory goes through one of the two corners, where the mean fitness is lowest.
Such a peak shift is only possible in viscous populations (except for some very extreme conditions). A viscous population is simulated in the Altruist software package as a population divided into small demes with a limited rate of migration between neighbor demes.
The simulations show the following evolutionary path:
One of the new genes, say a1, is created by mutation, and spreads by genetic drift. If the selection against the hybrid A1B0 is weak, then the a1 gene may become frequent, or even fixated, in a deme or a cluster of demes. The selection against the unfit hybrids A1B0 is relaxed in such a cluster, and hence such clusters can survive for many generations. Averaged over time and space, most hybrids are found in such clusters, even though the probability of the formation of a cluster of unfit hybrids is very low.
The next step (step 2 in Wright's shifting balance theory) is that the gene b1 is introduced into a cluster of A1B0 hybrids, either by mutation or by migration from a complementary A0B1 cluster, and a highly fit A1B1 individual is created. Under certain conditions, the A1B1 individuals will spread to other demes and cause fixation of the genes a1 and b1 (step 3).
If selection against the hybrids is stronger then the clusters of hybrids tend to be smaller and more short-lived, and the fraction of hybrids that are found in clusters becomes smaller. The formation of the first A1B1 individual may then take place without the formation of clusters, but the probability is extremely small because the a0 → a1 and b0 → b1 mutations have to take place close to each other in time and space.
A likely further step is that other genes in the genome, which previously were adapted to the A0B0 peak, are modified as an adaptation to the A1B1 peak. This fine-tunes the fitness further and makes the reverse path impossible, even if altered environmental conditions should again favor the A0B0 peak. This mechanism may account for the formation of new species.
|
|
| This figure shows a cluster of demes containing a1 genes. Blue squares
represent demes without a1 genes. Cyan, green, yellow, and
red squares represent demes with increasing frequencies of a1
genes. The first A1B1 phenotype has just appeared in
the yellow deme marked with an X. The b1 gene originates
from a mutation that happened within the same deme in the preceding
generation. (parameters: The figure is a section of a simulated population consisting of 1000 demes each containing 10 individuals on average. Selection against the A1B0 hybrids is 0.1%. Genes a1 and b1 are dominant. Migration rate is 10%) |
A more detailed study will be published later.
Barton, N H; Rouhani, S (1987): The Frequency of Shifts Between Alternative Equilibria. Journal of Theoretical Biology, vol. 125, no. 4, pp. 397-418.
